7 укажите что является результатом двойного оплодотворения. Опыление и двойное оплодотворение цветковых растений. Строение семязачатка. Двойное оплодотворение у покрытосеменных растений
Оплодотворение - это процесс слияния двух клеток, в результате чего происходит образование новой клетки, дающей начало другому организму этого же рода или вида. Что такое у цветковыхрастений и как оно происходит, читайте в данной статье.
Сущность оплодотворения
Оно происходит в результате слияния двух клеток, женской и мужской, и возникновения диплоидной зиготы. В каждой паре хромосом присутствует одна отцовская и одна материнская клетка. Сущность процесса оплодотворения заключается в том, чтобы восстановить и объединить наследственный материал родителей. Их потомство будет более жизнеспособным, так как соединит в себе самые полезные качества от отца и матери.
Оплодотворение - что такое?
Это процесс побуждения яйца к развитию в результате объединения ядер. Оплодотворение - что такое? Это необратимый процесс, который происходит в результате слияния разнополых гамет и объединения их ядер. не подвергается этой процедуре второй раз.
Но существуют растения, которые воспроизводят новое поколение только при помощи женской гаметы без оплодотворения. Такое размножение называется девственным. Примечательно, что эти два способа размножения у одного вида растений могут чередоваться.
Двойное оплодотворение цветковых растений
Обоих начал называются гаметами. Причем женскими являются яйцеклетки, а мужскими - спермии, которые у растений семенных неподвижные, а у споровых - подвижные. Оплодотворение - что такое? Это появление особой клетки - зиготы, содержащей наследственные признаки спермия и яйцеклетки.
Обладают сложным оплодотворением, которое называется двойным, поскольку, кроме яйцеклетки, оплодотворяется еще одна особая клетка. Формирование спермий происходит в пылинках пыльцы, а их созревание осуществляется в тычинках, точнее в их пыльниках. Местом образования яйцеклеток являются семязачатки, расположенные в завязи пестика. Когда яйцеклетка оплодотворится спермием, из семязачатка начинают развиваться семена.
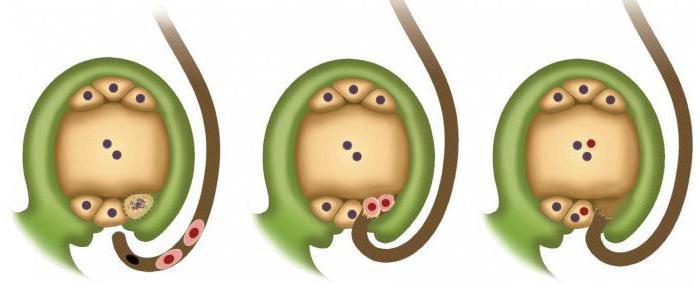
Чтобы оплодотворение у цветковых произошло, сначала нужно опылить растение, то есть на рыльце пестика должны попасть пылинки пыльцы. Оказавшись на рыльце, они начинают прорастать внутрь завязи, в результате чего образуется пыльцевая трубка. Одновременно с этим в пылинке происходит образование двух спермиев. Они не стоят на месте, а начинают продвигаться к пыльцевой трубке, которая проникает в семязачаток. Здесь в результате деления и удлинения одной клетки происходит образование зародышевого мешка.
Он нужен для расположения в нем яйцеклетки и еще одной клетки, в которой сосредоточен двойной набор наследственной информации. После этого происходит прорастание пыльцевой трубки в зародышевый мешок и слияние одного спермия с яйцеклеткой, в результате которого образуется зигота, а другого - с клеткой особой. Развитие зародыша происходит из зиготы. Второе слияние образует питательную ткань, или эндосперм, необходимый для питания зародыша в период роста.
Что нужно для существования каждого вида растений?
- Прежде всего необходимо восстановить диплоидный набор хромосом, а в его пределах - их парность.
- Обеспечить материальную непрерывность между поколениями, следующими чередой.
- Объединить в одном виде или роде наследственные свойства двух родителей.

Все это осуществляется на генетическом уровне. Для того чтобы оплодотворение осуществилось, созревание материнских и отцовских гамет должно произойти одновременно.
Оплодотворение у покрытосеменных растений
Этот процесс впервые охарактеризовал немецкий ученый Страсбургер во второй половине девятнадцатого века. Оплодотворение покрытосеменных растений происходит в результате слияния двух ядер разных гамет: с мужским и женским началом. Их цитоплазма не участвует в оплодотворении. Собственно оплодотворение происходит тогда, когда спермий сливается с ядром яйцеклетки.
Местом возникновения спермиев является пыльцевое зерно или пыльцевая труба. Зерно начинает прорастать после того, как попадает на рыльце. Время начала этого процесса у каждого растения разное, как и время оплодотворения. Например, пыльцевые зерна свеклы прорастают через два часа, а кукурузы - моментально. Первый признак прорастания зерна - его увеличение в объеме. Обычно одно пыльцевое зерно образует одну трубку. Но некоторые растения не подчиняются этому правилу и образуют несколько трубок, из которых только одна достигает своего развития.
Пыльцевая трубка с передвигающимися по ней спермиями растет и в конце концов разрывается. Все ее содержимое оказывается внутри зародышевого мешка. Один из проникших сюда спермиев внедряется в яйцеклетку и сливается с ее гаплоидным ядром. Оплодотворение - что такое? Это слияние двух ядер: спермия и яйцеклетки. Оплодотворенная яйцеклетка начинает делиться, получаются две новые клетки. Они делятся на четыре и так далее. Таким образом, происходит многократное деление, в результате которого развивается зародыш растения.

Покрытосеменные растения после процесса оплодотворения обладают способностью развивать дополнительный орган, который называется эндоспермом. Это не что иное, как питательная среда зародыша. При слиянии второго спермия и диплоидного ядра происходит образование определенного набора хромосом, из которых два - материнского происхождения, и один - отцовского. Таким образом, двойное оплодотворение организмов растительного происхождения осуществляется тогда, когда один спермий сливается с яйцеклеткой, а другой - с ядром клетки, расположенной в центре.
Отличительные черты покрытосеменных растений
- Большая приспособленность к произрастанию в разных условиях.
- Двойное оплодотворение, позволяющее иметь запас веществ, необходимых для нормального прорастания семян.
- Наличие триплоидного эндосперма.
- Образование семяпочек внутри завязи, при котором стенки пестика защищают их от повреждений.
- Развитие плода покрытосеменных растений из завязи.
- Нахождение семени внутри плода, стенки которого являются его защитой.
- Наличие цветка дает возможность насекомым.
Благодаря перечисленным признакам занимают господствующее положение в мире.
Особенность оплодотворения покрытосеменных растений
Она вытекает из того, что эти растения имеют двойное оплодотворение. Уникальная особенность представлена явлением, называемым ксениями. Его смысл заключается в том, что пыльца напрямую влияет на свойства и признаки эндосперма. Для примера возьмем кукурузу.

Она бывает с желтыми и белыми семенами. Их цвет зависит от оттенка эндосперма. При опылении женских цветков белозерной кукурузы пыльцой желтозерного сорта ее окраска все равно будет желтой, хотя развитие эндосперма происходит на растении с белыми зернами.
Какую роль играют цветковые растения?
Эти растения насчитывают 13 000 родов и 250 000 видов. Они получили широкое распространение по всему миру. Цветковые растения - это ключевые компоненты биосферы, производящие органические вещества, связывающие углекислоту и выделяющие кислород. Пастбищные цепи питания начинаются именно с них. Многие разновидности цветковых растений человек использует в пищу. Из них строят жилища и изготавливают различные хозяйственные материалы.

Не обходится без них и медицина. Отдельные виды покрытосеменных растений являются господствующими на планете, им отводится решающая роль в формировании покрова растительности и создании основной части наземной фитомассы. В конечном итоге именно этими растениями определяется возможность самого существования человека на земле как вида биологического.
Двойное оплодотворение
половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка (См. Зародышевый мешок). Д. о. открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия (См. Спермии) сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается Зародыш ,
из центральной клетки - Эндосперм . В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид (См. Синергиды),
которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке. При Д. о. ядра зародышевого мешка находятся в интерфазе (См. Интерфаза) и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать.
У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер. По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа Д. о.: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу (См. Профаза),
в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности (См. Плоидность);
у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие Д. о. - Ксении -
проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в Д. о., спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки. Лит.:
Навашин С. Г., Избр. труды, т. 1, М.- Л., 1951; Магешвар и П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Поддубная Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Steffen К., Fertilisation, в кн.: Maheshwari P. (ed.). Recent advances in the embryology of angiosperms, Delhi, 1963. И. Д. Романов.
Большая советская энциклопедия. - М.: Советская энциклопедия . 1969-1978 .
Смотреть что такое "Двойное оплодотворение" в других словарях:
Свойственно только цветковым растениям. При двойном оплодотворении один из спермиев сливается с яйцеклеткой, а второй с центральной клеткой зародышевого мешка. Из оплодотворенной яйцеклетки развивается зародыш, из центральной клетки вторичный… … Большой Энциклопедический словарь
Тип полового процесса, свойственный только цветковым растениям. Открыто в 1898 С. Г. Навашиным у лилейных. Д. о. заключается в том, что при формировании семени оплодотворяется не только яйцеклетка, но и центр, ядро зародышевого мешка. Из зиготы… …
двойное оплодотворение - Тип полового процесса, характерный для цветковых растений: один из спермиев оплодотворяет яйцеклетку, а другой (из той же пыльцевой трубки) оплодотворяет центральное ядро зародышевого мешка, в результате первого процесса образуется диплоидная… … Справочник технического переводчика
Свойственно только цветковым растениям. При двойном оплодотворении один из спермиев сливается с яйцеклеткой, а второй с центральной клеткой зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки вторичный… … Энциклопедический словарь
Double fertilization двойное оплодотворение. Тип полового процесса, характерный для цветковых растений: один из спермиев оплодотворяет яйцеклетку, а другой (из той же пыльцевой трубки
Свойственно только цветковым р ниям. При Д. о. один из спермиев сливается с яйцеклеткой, а второй с центр. клеткой зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центр. клетки вторичный эндосперм семени, содержащий… … Естествознание. Энциклопедический словарь
двойное оплодотворение - процесс оплодотворения, происходящий у покрытосеменных растений, в котором принимают участие оба образующихся спермия. Один из них сливается с яйцеклеткой, второй – с центральной диплоидной клеткой зародышевого мешка. Открыто С. Г. Навашиным в… … Анатомия и морфология растений
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ - половой процесс у покрытосеменных, заключающийся в слиянии одной мужской гаметы пыльценой трубки (спермия) с яйцеклеткой зародышевого мешка, а второй мужской гаметы с вторичным ядром зародышевого мешка … Словарь ботанических терминов
двойное оплодотворение по навашину - ЭМБРИОЛОГИЯ РАСТЕНИЙ ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ ПО НАВАШИНУ – слияние яйцеклетки и спермия с образованием зиготы (2п) и одновременное слияние другого спермия и двойного ядра с образованием первичного ядра эндосперма (3п). Характерная особенность всех … Общая эмбриология: Терминологический словарь
Сингамия, слияние мужской половой клетки (сперматозоид, спермий) с женской (яйцо, яйцеклетка), приводящее к образованию зиготы, края даёт начало новому организму. Уживотных О. предшествует осеменение. В процессе О. осуществляются активация яйца,… … Биологический энциклопедический словарь
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая - гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на "мега" (или "макро"). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов - материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна - отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап - микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки - спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими - синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.

Особенности пыльцевой трубки
Двойное оплодотворение сопровождается взаимодействием ее с тканями спорофита. Оно достаточно специфично. Этот процесс регулируется активностью химических соединений. Установлено, что если пыльцу промыть в дистиллированной воде, она потеряет способность к прорастанию. Если же полученный раствор сконцентрировать, а затем ее обработать, то она снова станет полноценной. Развитие пыльцевой трубки после прорастания контролируют ткани пестика. К примеру, у хлопчатника ее рост до яйцеклетки занимает порядка 12-18-ти часов. Однако уже спустя 6 часов вполне можно определить, к какой именно семяпочке будет направляться пыльцевая трубка. Это понятно потому, что в ней начинается разрушение синергиды. В настоящее время не установлено, как растение может направить развитие трубки в нужном направлении и каким образом о приближении узнает синергида.

"Запрет" на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. "Запрет" на самоопыление проявляется в том, что спорофит "идентифицирует" собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же - короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца - мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.

Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев "самонесовместимость" пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, - s1s2, а вырабатывающего пыльцу - s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
В процессе эволюции растительного мира у цветковых растений (и только у них) появилось такое явление как двойное оплодотворение, в результате которого образуется семя. У голосеменных растений также образуются семена, но двойного оплодотворения нет. Оплодотворению предшествует опыление, т. е. перенос пыльцы с тычинок одного цветка на пестик чаще всего другого цветка. При двойном оплодотворении в семязачаток проникают два спермия, один из которых сливается с яйцеклеткой, а второй - с крупной центральной клеткой.
Пыльцевые зерна разных цветковых растений имеют различную форму. При этом чаще всего поверхность пыльцевых зерен шероховатая, что позволяет им удерживаться на теле насекомых-опылителей и потом на рыльце пестика. Кроме того, рыльцем выделяется липкая жидкость, удерживающая пыльцу. На рыльце пестика пыльцевое зерно образует пыльцевую трубку , которая растет между клетками рыльца и столбика пестика, после чего врастает в полость завязи пестика.
В полости завязи может находиться один семязачаток, несколько или множество. Их количество зависит от вида растения. Семязачатки по-другому называются семяпочками . Если в завязи несколько семязачатков, то каждый из них опыляется своим пыльцевым зерном (содержащимися в нем спермиями), т. е. в таком случае через пестик будет прорастать несколько пыльцевых трубок.
Семязачатки отрастают от внутренней поверхности стенок завязи в полость завязи. Семязачаток состоит из покрова и ткани центральной части, где образуются восемь гаплоидных клеток (имеющих одинарный набор хромосом). Две из этих клеток сливаются, в результате образуется крупная центральная клетка , у которой восстанавливается двойной набор хромосом.
У семязачатка со стороны, противоположной месту прикрепления к завязи, находится пыльцевход , представляющий собой небольшое отверстие, ведущее к центральной части семязачатка.
В кончике растущей пыльцевой трубки находятся два спермия . У спермиев в отличие от сперматозоидов нет хвостика, и поэтому спермии неподвижны. Когда трубка врастает в семязачаток через пыльцевход, то один спермий сливается с одной из гаплоидных клеток, которая играет роль яйцеклетки . В результате этого оплодотворения образуется зигота с двойным набором хромосом. В последствии из нее развивается зародыш семени .
Второй спермий сливается с центральной клеткой. В результате этого оплодотворения в последствии образуется так называемый эндосперм . Для него характерен тройной набор хромосом, что уникально, так как клетки тела покрытосеменных и многих других организмов имеют двойной набор хромосом.
Эндосперм представляет собой ткань, содержащую запас питательных веществ. Эти вещества зародыш использует в процессе развития семени или при прорастании семени. В первом случае вместо эндосперма в зрелом семени основную массу занимают органы зародыша (чаще всего крупные семядоли), во втором случае - эндосперм остается.
При созревании семени покровы семязачатка превращаются в семенную кожуру .
Водоросли.Общая характеристика. Водоросли – растения, живущие преимущественно в воде. Тело их не расчленено на органы и ткани. Органы размножения одноклеточные. Это одни из древнейших представителей растительного мира. Водоросли бывают одноклеточными (хламидоионада, хлорелла), колониальные (носток) и многоклеточные (спирогира, ламинария,). Цитологические особенности:
Клетки покрыты клеточной стенкой. Характерно наличие хроматофоров , носителей окраски. Хроматофоры – органеллы, в которых происходит фотосинтез. Окраска зависит от глубины обитания водорослей: на большой глубине – буро-красные, ближе к поверхности – зеленые. В хроматофорах всех водорослей содержатся пиреноиды , они синтезируют крахмал.
Одноклеточные формы могут быть подвижными (со жгутиками) и неподвижными.
Также водоросли бывают прикрепленные (бентосные) и свободноплавающие (планктонные) Размножение: Вегетативное – новые водоросли образуются из обрывков нитей, кусков слоевищ и т.д. Бесполое – содержимое одной клетки (зооспорангия) делится многократно, образую новые подвижные клетки (зооспоры). Каждая из них дает начало новой особи. Половое размножение – широко распространено у водорослей. Формы полового процесса разнообразны: изогамия (♂ подвижна, ♀ подвижна, одинаковые по размеру), гетерогамия (♂ подвижна, ♀ подвижна, ♀ больше, чем ♂), оогамия (♂ подвижна, ♀ неподвижна, ♀ больше, чем ♂), конъюгация (сливается содержимое двух вегетативных клеток). Образовавшаяся зигота покрывается толстой клеточной стенкой, накапливает запасные питательные вещества и в состоянии покоя способна легка переносить неблагоприятные условия.
1 .Общая характеристика отдела Папоротниковые . За исключением нескольких родов все папоротники - равноспоровые. У них наблюдается смена поколений с преобладание спорофита над гаметофитом. Спорофит представлен многолетними корневищными травами с крупными, обычно перисто-рассеченными листьями, на нижней стороне которых расположены спорангии. В условия тропического климата встречаются древовидные папоротники. Листья папоротников нарастают верхушкой. То, что у папоротника напоминает лист - вовсе не лист, а по своей природе - целая система ветвей, да ещё расположенных в одной плоскости. Так это и называется - плосковетка, или вайя , или, ещё одно название, - предпобег. В спорангиях в результате мейоза образуются споры (n). Спорангии лопаются, споры высыпаются, прорастает заросток (гаметофит). Заростки представляют собой сердцевидную зеленую пластинку. Точка роста находится в выемке. От нижней стороны отходят ризоиды. На заростке образуются антеридии (в них образуются сперматозоиды) и архегонии (в ней образуется яйцеклетка). Во время дождя или обильной росы сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку. Из зиготы развивается зародыш, затем взрослый спорофит. Современные папоротники насчитывают около 300 родов и 12 000 видов. Основные представители: Щитовник мужской, Страусник обыкновенный, Сальвиния плавающая (разноспоровая)

Опыление у покрытосемянных растений. Приспособление цветков к различным видам опыления. Микро- и макроспорогенез у растений Опыление – перенос пыльцы с тычинок на рыльца пестика. Различают самоопыление и перекрестное опыление. Самоопыление: пыльца опыляет рыльце пестика того же цветка (пшеница, ячмень, овес, просо, горох, фасоль, бобы, хлопчатник, лен, томат и др.) . Преобладает у 10% растений. Самоопыление происходит как у раскрывшихся цветков: сельдерейные, так и у закрытых: арахис, фиалка. Однако для эволюционного развития этот вид самоопыления не совершенен, т.к.не несет новых генетических признаков. Поэтому многие растения имеют приспособления, препятствующие самоопылению: Двудомность (♂ и ♀ цветки формируются на разных растениях) Однодомность (♂ и ♀ цветки формируются на одном растении, но в разных цветках) Дихогамия – разновременное созревание пыльцы и рыльца Гетеростилия – рыльца и тычиночные нити различаются по длине Самонесовместимость. Перекрестное опыление : пыльца опыляет рыльца других цветков. Различают 2 типа: гейтоногамия – опыление в пределах одного растения ксеногамия – опыление в пределах разных цветков (наиболее оптимально для эволюционного процесса) Различают несколько видов: Абиотическое – при помощи неживых факторов внешней среды Анемофилия (ветром)
Гидрофилия (водой)
Биотическое – с помощью животных.
Энтомофилия – опыление насекомыми Орнитофилия – опыление мелкими птицами (колибри)

Строение семязачатка. Двойное оплодотворение у покрытосеменных растений.
Строение семязачатка. Семязачаток состоит из центральной части - нуцеллуса и окружающих его одного или двух покровов - интегументов, которые над верхушкой нуцеллуса образуют небольшой канал - микропиле. Семязачаток сообщается с плацентой с помощью семяножки, или фуникулуса. Нуцеллус является аналогом макроспорангия, в котором развивается одна макроспора.Двойное оплодотворение было открыто С.Г.Навашиным в 1898 г. Оплодотворение у покрытосемянных принято называть двойным, т.к. оба спермия сливаются с клетками зародышевого мешка. Один сливается с яйцеклеткой, в результате образуется зигота. Второй сливается с центральным ядром, в результате образуется триплоидная клетка (3n). Прочие клетки зародышевого мешка дегенерируют. После двойного оплодотворения из зиготы развивается зародыш, а из триплойдной клетки – эндосперм (питательная ткань), из нуцеллуса образуется перисперм (дополнительная питательная ткань), из интегумента – семенная кожура, из семязачатка – семя, из завязи – плод. Преимущества двойного оплодотворения заключаются в том, что одновременно с зиготой формируется триплоидная клетка (3n), которая делится быстрее, чем зигота. Соответственно, эндосперм образуется быстрее, чем растет зародыш. Поэтому не нужно запасать питательные вещества до оплодотворения, в отличие от голосеменных, у которых довольно мощный гаплоидный эндосперм развивается до оплодотворения. Этим достигается существенная экономии я энергетических ресурсов организма. Семязачатки покрытосеменных, не обремененные запасающей питательной тканью впрок, развиваются гораздо быстрее, чем у голосеменных.


1 - покровы семязачатка, или интегументы (а - наружный, б - внутренний), 2 - микропиле, 3 - халаза, 4 - фуникулус, 5 - нуцеллус, 6 -зародышевый мешок, 7 - яйцеклетка, 8 - синергиды, 9 - антиподы, 10 - вторичное ядро, 11 - плацента, 12 - проводящий пучок. После оплодотворения из зиготы развивается зародыш, из центрального ядра – эндосперм, из нуцеллуса – перисперм, из интигументов – семенная кожура, из семязачатка – семя, из завязи – плод.
Высшие растения. Общая характеристика и цикл развития.
Высшие растения , или Наземные растения ,- тип зелёных растений, которым свойственна дифференциация тканей, в отличие от низших растений - водорослей. К высшим растениям относятся мхи и сосудистые растения (папоротникообразные, псилотовые,хвощевидные, плауновидные, голосеменные и покрытосеменные).
Развитие специализированных тканей было важным условием для выхода растений на сушу. Для комфортного существования в воздушной среде растениям было необходимо развить как минимум эпидермис с устьицами для защиты от высыхания и теплообмена и проводящие ткани для обмена минеральных и органических веществ. Результатом выхода растений на сушу также стало разделение организма растения на корень, стебель и лист.
В ЖЦ высших растений происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено спорофитом (2n), половое – гаметофитом (n). Спорофит – это растение, образующее споры. В многоклеточных спорангиях в результате мейотического деления формируются споры (n). Растения, у которых все споры одинаковые – равноспоровые, у более высокоорганизованных споры разной величины (микроспоры и мегаспоры) – это разноспоровые растения. Гаметофит – растение, образующее гаметы. Гаметы равиваются в многоклеточных органах полового размножения: яйцеклетки – в архегониях, сперматозойды – в антеридиях. Гаметофит вырастает из споры. У равноспоровых растений гаметофит обоеполый, у разноспоровых – однополый. В результате оплодотворения образуется зигота, из которой прорастает новый спорофит. Все высшие растения делятся на 2 группы по доминированию в ЖЦ гаметофита и спорофита: Растения с доминирующим гаметофитом – отдел Моховидные Растения с доминирующим спорофитом – все остальные
В целом для эволюции высших растений характерна тенденция к усложнению и усовершенствованию спорофита при одновременной редукции гаметофита.

Моховидные. Общая характеристика. Цикл развития мха Кукушкин лен. В отдел входит более 25 000 видов сравнительно просто организованных травянистых растений. В цикле развития преобладает гаметофит. У более примитивных форм он представлен талломом, а у остальных – расчленен не стебель и листья. Корней нет, из заменяют ризоиды. Спорофит самостоятельно не существует, развивается на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий. Отдел делится на 3 класса: Антоцеротовые, Печеночные, Листостебельные мхи. Наиболее известен вид – Кукушкин лен (Класс Листостебельные мхи). Представляет собой прямостоячие стебли (15-20 см) густо покрытые жесткими острыми листьями. Прикрепляется к земле ризоидами. Гаметофиты раздельнополые. На верхушке мужских особей развиваются антеридии, окруженные красно-бурыми листьями (n), на верхушках женских – архегонии (n). Оплодотворение происходит подвижными двужгутиковыми сперматозоидами во влажную погоду. Из зиготы на верхушке женского гаметофита развивается спорофит (2n), имеющий вид коробочки на длинной ножке. Внутри коробочки – спорангий, где после мейоза образуются споры (n). После образования спор колпачок, а затем крышечка отделяются и споры высыпаются. Из споры сначала образуется протонема, на которой из особых почек формируются листостебельные побеги – гаплоидное поколение (n).



Общая хар-ка отдела Плауновидные. Цикл развития Плауна булавовидного. Плауновидные – очень древняя группа, возникшая в силуре, а в каменноугольном периоде достигла своего расцвета. Это были огромные деревья, образующие целые леса. В современной флоре они представлены вечнозелеными многолетними травами, реже полукустарниками. Около 1000 видов сохранилось. Плауновидные имеют мелкие листья с одной жилкой микрофильного типа. Выделяют 2 класса: равноспоровые Плауновые и разноспоровые Полушниковые. ^ Плаун булавовидный . В цикле развития преобладает спорофит (2n). Он представляет собой длинные стелющиеся ветвящиеся побеги густо усажены жесткими мелкими листьями. От стебля отходят тонкие придаточные корни. В середине лета на верхушках появляются спороносные колоски. Колосок состоит из оси и сидящих на ней листочков (спорофиллов). На верхней стороне спорофиллов находится спорангий на короткой ножке. В результате мейоза в нем образуются споры (n). Из спор развивается заросток (гаметофит) (n). Заросток развивается под землей. От нижней его части отходят ризоиды. Через них он врастает в грибы, образуя микоризу. Живет в симбиозе с грибом, питается от него. Растет очень долго (15-20 лет). На верхней стороне его образуется антеридий и архегонии. Двужгутиковый сперматозоид выходи из антеридия, проникает в архегоний и оплодотворяет яйцеклетку. В результате образуется зигота, из которой развивается новый спорофит.





