Schwankungen in der Anzahl der Raubtiere und Beutetiere. Schwingungen des Räuber-Beute-Systems (Lotka-Voltaire-Modell). Simulationsmodellierung des „Predator-Prey“-Systems
zur Vereinbarung vom ___.___, 20___ über die Erbringung bezahlter Bildungsdienstleistungen
Ministerium für Bildung und Wissenschaft Russische Föderation
Lysvensky-Filiale
Staatliche Technische Universität Perm
Fakultät für Wirtschaftswissenschaften
in der Disziplin „Systemmodellierung“
Thema: Raubtier-Beutesystem
Vollendet:
Studentengr. BIVT-06
------------------
Vom Lehrer überprüft:
Schestakow A. P.
Lysva, 2010
Aufsatz
Unter Raub versteht man eine trophische Beziehung zwischen Organismen, bei der einer von ihnen (das Raubtier) den anderen (die Beute) angreift und sich von Teilen seines Körpers ernährt, d. h., das Opfer wird normalerweise getötet. Raubtiere stehen im Gegensatz zum Verzehr von Leichen (Nekrophagie) und organischen Zersetzungsprodukten (Detritophagie).
Sehr beliebt ist auch eine andere Definition von Raubtier, die vorschlägt, dass nur Organismen, die Tiere fressen, als Raubtiere bezeichnet werden, im Gegensatz zu Pflanzenfressern, die Pflanzen fressen.
Als Fressfeinde können neben vielzelligen Tieren auch Protisten, Pilze und höhere Pflanzen fungieren.
Die Populationsgröße von Raubtieren beeinflusst die Populationsgröße ihrer Beute und umgekehrt. Die Populationsdynamik wird durch das mathematische Lotka-Volterra-Modell beschrieben. Dieses Modell stellt jedoch einen hohen Abstraktionsgrad dar und beschreibt nicht die tatsächliche Beziehung zwischen Raubtier und Beute. und kann nur als erster Grad der Annäherung an die mathematische Abstraktion betrachtet werden.
Im Prozess der Koevolution passen sich Räuber und Beute aneinander an. Raubtiere tauchen auf und entwickeln Mittel zur Erkennung und zum Angriff, und Opfer verfügen über Mittel zur Geheimhaltung und Verteidigung. Daher kann den Opfern der größte Schaden durch für sie neue Raubtiere zugefügt werden, mit denen sie noch kein „Wettrüsten“ begonnen haben.
Raubtiere können sich auf eine oder mehrere Beutearten spezialisieren, was sie im Durchschnitt erfolgreicher bei der Jagd macht, aber auch ihre Abhängigkeit von diesen Arten erhöht.
Raubtier-Beute-System.
Die Interaktion zwischen Raubtieren und Beutetieren ist die Hauptform der vertikalen Beziehung zwischen Organismen, bei der Materie und Energie über Nahrungsketten übertragen werden.
Gleichgewicht von V. x. - Und. lässt sich am einfachsten erreichen, wenn es mindestens drei Glieder in der Nahrungskette gibt (z. B. Gras – Wühlmaus – Fuchs). Gleichzeitig wird die Dichte der Phytophagenpopulation durch Beziehungen sowohl zum unteren als auch zum oberen Glied der Nahrungskette reguliert.
Abhängig von der Art der Beute und der Art des Raubtiers (True, Grasfresser) sind unterschiedliche Abhängigkeiten von der Dynamik ihrer Populationen möglich. Darüber hinaus wird das Bild durch die Tatsache verkompliziert, dass Raubtiere sehr selten monophag sind (d. h. sich von einer Beuteart ernähren). Meistens dann, wenn die Population einer Beuteart erschöpft ist und deren Gewinnung zu viel erfordert hohe Kosten Kräfte wechseln Raubtiere zu anderen Beutearten. Darüber hinaus kann eine Beutepopulation von mehreren Raubtierarten ausgebeutet werden.
Aus diesem Grund kommt der in der Umweltliteratur häufig beschriebene Effekt einer pulsierenden Beutepopulationsgröße, gefolgt von einer mit einer gewissen Verzögerung pulsierenden Populationsgröße des Raubtiers, in der Natur äußerst selten vor.
Das Gleichgewicht zwischen Raubtieren und Beutetieren wird bei Tieren durch spezielle Mechanismen aufrechterhalten, die eine vollständige Ausrottung der Opfer verhindern. So können Opfer:
- vor einem Raubtier davonlaufen (in diesem Fall erhöht sich durch den Wettbewerb die Mobilität von Beute und Raubtieren, was besonders typisch für Steppentiere ist, die sich nirgendwo vor ihren Verfolgern verstecken können);
- eine schützende Farbe erhalten (<притворяться>Blätter oder Zweige) oder im Gegenteil eine helle (z. B. rote) Farbe, die das Raubtier vor dem bitteren Geschmack warnt;
- in Notunterkünften verstecken;
- Gehen Sie zu aktiven Abwehrmaßnahmen über (gehörnte Pflanzenfresser, stachelige Fische), oft gemeinsam (Beutevögel vertreiben gemeinsam den Drachen, männliche Hirsche und Saigas besetzen).<круговую оборону>von Wölfen usw.).
Zurück in den 20er Jahren. A. Lotka und etwas später, unabhängig von ihm, V. Volterra schlugen mathematische Modelle vor, die konjugierte Schwankungen in den Populationen von Raubtieren und Beutetieren beschreiben. Betrachten wir die einfachste Version des Lotka-Volterra-Modells. Das Modell basiert auf einer Reihe von Annahmen:
1) Die Beutepopulation wächst in Abwesenheit eines Raubtiers exponentiell,
2) der Druck von Raubtieren hemmt dieses Wachstum,
3) Die Sterblichkeit der Beute ist proportional zur Häufigkeit der Begegnungen zwischen Raubtier und Beute (oder andernfalls proportional zum Produkt ihrer Populationsdichten);
4) Die Geburtenrate eines Raubtiers hängt von der Intensität des Beutekonsums ab.
Die momentane Änderungsrate der Beutepopulationsgröße kann durch die Gleichung ausgedrückt werden
dN f /dt = r 1 N f - p 1 N f N x,
wo r 1 - spezifische momentane Rate des Populationswachstums der Beute, p 1 – Konstante, die die Sterblichkeit der Beute mit der Dichte des Raubtiers verbindet, a N Und Nx - Dichte der Beute bzw. des Raubtiers.
Es wird angenommen, dass die momentane Wachstumsrate der Raubtierpopulation in diesem Modell der Differenz zwischen der Geburtenrate und der konstanten Sterblichkeit entspricht:
dN x /dt = p 2 N x N x – d 2 N x,
wo p 2 - Konstante, die die Geburtenrate in einer Raubtierpopulation mit der Beutedichte in Beziehung setzt, a d 2 - spezifische Sterblichkeitsrate des Raubtiers.
Gemäß den obigen Gleichungen wird jede der interagierenden Populationen in ihrem Wachstum nur durch die andere Population begrenzt, d. h. Der Anstieg der Zahl der Opfer wird durch den Druck der Raubtiere begrenzt, und der Anstieg der Zahl der Raubtiere wird durch die unzureichende Zahl der Opfer begrenzt. Es wird keine Selbstlimitierung der Populationen angenommen. Man geht beispielsweise davon aus, dass immer genug Nahrung für das Opfer vorhanden ist. Es ist auch nicht zu erwarten, dass die Beutepopulation der Kontrolle des Raubtiers entgeht, obwohl dies tatsächlich recht häufig vorkommt.
Trotz aller Konventionalität des Lotka-Volterra-Modells verdient es Aufmerksamkeit, schon allein deshalb, weil es zeigt, wie selbst ein solch idealisiertes System der Interaktion zwischen zwei Populationen zu einer recht komplexen Dynamik ihrer Zahlen führen kann. Die Lösung des Systems dieser Gleichungen ermöglicht es uns, die Bedingungen für die Aufrechterhaltung einer konstanten (Gleichgewichts-)Anzahl jeder Art zu formulieren. Die Beutepopulation bleibt konstant, wenn die Dichte des Raubtiers gleich r 1 /p 1 ist, und damit die Raubtierpopulation konstant bleibt, muss die Dichte der Beute gleich d 2 /p 2 sein. Tragen wir die Opferdichte auf der x-Achse ein N Und , und entlang der Ordinate - die Dichte des Raubtiers N X, dann sind die Isoklinen, die den Zustand der Konstanz von Raubtier und Beute zeigen, zwei gerade Linien senkrecht zueinander und zu den Koordinatenachsen (Abb. 6, a). Es wird davon ausgegangen, dass unterhalb einer bestimmten Beutedichte (gleich d 2 /p 2) die Dichte des Raubtiers immer abnimmt und darüber immer zunimmt. Dementsprechend nimmt die Dichte der Beute zu, wenn die Dichte des Raubtiers unter dem Wert gleich r 1 / p 1 liegt, und nimmt ab, wenn sie über diesem Wert liegt. Der Schnittpunkt der Isoklinen entspricht der Bedingung einer konstanten Häufigkeit von Raubtieren und Beutetieren, und andere Punkte auf der Ebene dieses Diagramms bewegen sich entlang geschlossener Trajektorien und spiegeln somit regelmäßige Schwankungen in der Häufigkeit von Raubtieren und Beutetieren wider (Abb. 6, B). Der Schwingungsbereich wird durch das anfängliche Verhältnis der Dichten von Räuber und Beute bestimmt. Je näher es am Schnittpunkt der Isoklinen liegt, desto kleiner ist der durch die Vektoren beschriebene Kreis und desto kleiner ist dementsprechend die Schwingungsamplitude.
Reis. 6. Grafische Darstellung des Lotka-Voltaire-Modells für das Raubtier-Beute-System.
Einer der ersten Versuche, Schwankungen in der Häufigkeit von Raubtieren und Beutetieren in Laborexperimenten zu ermitteln, gehörte G.F. Gause. Die Objekte dieser Experimente waren die Paramecium ciliate (Paramecium caudatum) und das räuberische Wimpertier Didinium (Didinium Nasutum). Die Nahrung für Paramecium war eine Suspension von Bakterien, die dem Medium regelmäßig zugesetzt wurde, und Didinium ernährte sich ausschließlich von Paramecium. Dieses System erwies sich als äußerst instabil: Der Druck des Raubtiers führte mit zunehmender Zahl zur vollständigen Ausrottung der Opfer, woraufhin die Population des Raubtiers selbst ausstarb. Um die Experimente zu erschweren, richtete Gause einen Unterschlupf für das Opfer ein, indem er ein wenig Glaswolle in Reagenzgläser mit Ciliaten einführte. Paramecia konnte sich frei zwischen den Wattefäden bewegen, Didinium jedoch nicht. In dieser Version des Experiments fraß Didinium alle Paramecia, die im wattefreien Teil des Reagenzglases schwammen, und starb aus, und die Paramecia-Population wurde dann durch die Fortpflanzung von Individuen, die im Tierheim überlebten, wiederhergestellt. Gause konnte nur dann einen gewissen Anschein von Schwankungen in der Anzahl von Raubtieren und Beutetieren erzielen, wenn er von Zeit zu Zeit sowohl Beute als auch Raubtiere in die Kultur einführte und so eine Einwanderung simulierte.
40 Jahre nach Gauses Arbeit wurden seine Experimente von L. Luckinbill wiederholt, der Ciliaten als Opfer verwendete Paramecium Aurelia, und als Raubtier desselben Didinium Nasutum. Luckinbill gelang es, mehrere Schwankungszyklen in der Zahl dieser Populationen zu erreichen, allerdings nur in dem Fall, in dem die Dichte des Parameciums durch einen Mangel an Nahrung (Bakterien) begrenzt war und der Kulturflüssigkeit Methylzellulose zugesetzt wurde – eine Substanz, die die Zahl reduziert Geschwindigkeit der Bewegung von Raubtier und Beute und verringert daher die Häufigkeit ihrer möglichen Begegnungen. Es stellte sich auch heraus, dass es einfacher ist, Oszillationen zwischen Räuber und Beute zu erreichen, wenn das Volumen des Versuchsgefäßes vergrößert wird, obwohl auch in diesem Fall die Bedingung einer Nahrungsbeschränkung der Beute erforderlich ist. Wenn dem System aus Raubtier und Beute, die in einem oszillierenden Modus nebeneinander existieren, überschüssige Nahrung hinzugefügt wurde, dann war die Reaktion ein rascher Anstieg der Zahl der Beute, gefolgt von einem Anstieg der Zahl der Raubtiere, was wiederum zur vollständigen Ausrottung von Raubtieren führte die Beutepopulation.
Die Lotka- und Volterra-Modelle dienten als Anstoß für die Entwicklung einer Reihe weiterer realistischerer Modelle des Raubtier-Beute-Systems. Insbesondere ein recht einfaches grafisches Modell, das das Verhältnis verschiedener Beute-Isoklinen analysiert Raubtier, wurde von M. Rosenzweig und R. MacArthur (Rosenzweig, MacArthur) vorgeschlagen. Laut diesen Autoren sind stationäre ( = konstant) Die Anzahl der Beute in den Koordinatenachsen der Dichte von Raubtier und Beute kann in Form einer konvexen Isokline dargestellt werden (Abb. 7, a). Ein Punkt, an dem sich die Isokline mit der Beutedichtelinie schneidet, entspricht der minimal zulässigen Beutedichte (die Bevölkerung darunter ist einem sehr hohen Risiko des Aussterbens ausgesetzt, allein schon aufgrund der geringen Häufigkeit von Treffen zwischen Männern und Frauen), und der andere Punkt entspricht dem Maximum, bestimmt durch die Menge der verfügbaren Nahrung oder die Verhaltensmerkmale der Beute selbst. Wir möchten betonen, dass wir immer noch über minimale und maximale Dichten in Abwesenheit eines Raubtiers sprechen. Wenn ein Raubtier auftaucht und seine Zahl zunimmt, sollte die minimal zulässige Beutedichte natürlich höher und die maximale niedriger sein. Jeder Wert der Beutedichte muss einer bestimmten Räuberdichte entsprechen, bei der eine Konstanz der Beutepopulation erreicht wird. Die geometrische Lage solcher Punkte ist die Beute-Isokline in den Dichtekoordinaten von Räuber und Beute. Vektoren, die die Richtung der Änderung der Beutedichte (horizontal ausgerichtet) zeigen, haben auf verschiedenen Seiten der Isokline unterschiedliche Richtungen (Abb. 7a).

Reis. 7. Isokline stationärer Populationen von Beutetieren (a) und Raubtieren (b).
Für das Raubtier wurde ebenfalls eine Isokline in denselben Koordinaten konstruiert, die dem stationären Zustand seiner Population entspricht. Vektoren, die die Richtung der Änderung der Raubtierhäufigkeit anzeigen, sind nach oben oder unten ausgerichtet, je nachdem, auf welcher Seite der Isokline sie sich befinden. Die Form der Raubtier-Isokline, dargestellt in Abb. 7, B. wird erstens durch das Vorhandensein einer bestimmten Mindestdichte der Beute bestimmt, die ausreicht, um die Population des Raubtiers aufrechtzuerhalten (bei einer geringeren Dichte der Beute kann das Raubtier seine Zahl nicht erhöhen), und zweitens durch das Vorhandensein von a bestimmte maximale Dichte des Raubtiers selbst, oberhalb derer die Zahl unabhängig von der Häufigkeit der Opfer abnimmt.

Reis. 8. Die Entstehung von Oszillationsmodi im Räuber-Beute-System in Abhängigkeit vom Standort des Räubers und der Isoklinen der Beute.
Bei der Kombination von Beute- und Raubtier-Isoklinen in einem Diagramm sind drei verschiedene Optionen möglich (Abb. 8). Wenn die Räuber-Isokline die Beute-Isokline an der Stelle schneidet, an der sie bereits abnimmt (bei hoher Beutedichte), bilden die Vektoren, die Änderungen in der Häufigkeit von Räuber und Beute zeigen, eine nach innen gedrehte Flugbahn, was gedämpften Fluktuationen in der Häufigkeit entspricht von Beute und Raubtier (Abb. 8, A). Für den Fall, dass die Isokline des Raubtiers die Isokline der Beute in ihrem aufsteigenden Teil schneidet (d. h. im Bereich niedriger Werte der Beutedichte), bilden die Vektoren eine sich abwickelnde Flugbahn, und mit zunehmender Zunahme treten Schwankungen in der Anzahl der Raubtiere und Beute auf Amplitude bzw. (Abb. 8, B). Wenn die Isokline des Raubtiers die Isokline der Beute im Bereich ihrer Spitze schneidet, bilden die Vektoren einen geschlossenen Kreis und Schwankungen in der Anzahl der Beute und des Raubtiers sind durch eine stabile Amplitude und Periode gekennzeichnet (Abb. 8, V).
Mit anderen Worten entsprechen gedämpfte Schwingungen einer Situation, in der ein Raubtier einen spürbaren Einfluss auf eine Beutepopulation ausübt, die nur eine sehr hohe Dichte (nahe dem Maximum) erreicht hat, und es treten Schwingungen mit zunehmender Amplitude auf, wenn das Raubtier dazu in der Lage ist Auch bei geringer Beutedichte lässt sich die Zahl schnell erhöhen und so schnell vernichten. In anderen Versionen ihres Modells zeigten Posenzweig und McArthur, dass Räuber-Beute-Oszillationen durch die Einführung eines „Zufluchtsorts“, d. h. Dies deutet darauf hin, dass es in einem Gebiet mit geringer Beutedichte ein Gebiet gibt, in dem die Beutehäufigkeit unabhängig von der Anzahl der vorhandenen Raubtiere zunimmt.
Der Wunsch, Modelle durch Erhöhung ihrer Komplexität realistischer zu machen, manifestierte sich nicht nur in den Werken von Theoretikern, sondern auch Experimentatoren. Interessante Ergebnisse erzielte insbesondere Huffaker, der am Beispiel einer kleinen pflanzenfressenden Milbe die Möglichkeit der Koexistenz von Räuber und Beute im Oszillationsmodus aufzeigte Eotetranychus Sexmaculatus und eine räuberische Zecke greift ihn an Typhlodromus occidentalis. Als Nahrung für die pflanzenfressende Milbe dienten Orangen auf Tabletts mit Löchern (wie sie zum Aufbewahren und Transportieren von Eiern verwendet werden). Die Originalversion enthielt 40 Löcher auf einem Tablett, von denen einige Orangen (teilweise geschält) und andere Gummibälle enthielten. Beide Zeckenarten vermehren sich parthenogenetisch sehr schnell, sodass die Art ihrer Populationsdynamik in relativ kurzer Zeit aufgedeckt werden kann. Nachdem er 20 Weibchen der pflanzenfressenden Milbe auf ein Tablett gelegt hatte, beobachtete Huffaker das schnelle Wachstum seiner Population, die sich auf dem Niveau von 5-8.000 Individuen (pro Orange) stabilisierte. Wenn der wachsenden Beutepopulation mehrere Individuen eines Raubtiers hinzugefügt wurden, nahm die Population der Beutetiere schnell zu und starb aus, als alle Opfer gefressen waren.
Durch die Vergrößerung der Schale auf 120 Löcher, in denen einzelne Orangen zufällig auf viele Gummibälle verteilt waren, konnte Huffaker die Koexistenz von Raubtier und Beute verlängern. Wie sich herausstellte, spielt das Verhältnis ihrer Ausbreitungsraten eine wichtige Rolle bei der Interaktion zwischen Räuber und Beute. Huffaker schlug vor, dass durch die Erleichterung der Bewegung der Beute und die Erschwerung der Bewegung des Raubtiers die Zeit ihres Zusammenlebens verlängert werden könnte. Dazu wurden auf einem Tablett mit 120 Löchern 6 Orangen zufällig zwischen Gummibällen platziert, und um die Löcher mit Orangen wurden Vaseline-Barrieren gebaut, um die Ausbreitung des Raubtiers zu verhindern, und um die Ansiedlung der Beute zu erleichtern, wurden Holzpflöcke angebracht auf dem Tablett befestigt und dienen als eine Art „Startrampe“ für pflanzenfressende Milben (Tatsache ist, dass diese Art dünne Fäden produziert und mit deren Hilfe in der Luft schweben und sich mit dem Wind ausbreiten kann). In einem solch komplexen Lebensraum existierten Räuber und Beute acht Monate lang nebeneinander, was drei vollständige Zyklen von Populationsschwankungen zeigt. Die wichtigsten Bedingungen für dieses Zusammenleben sind folgende: Heterogenität des Lebensraums (im Sinne des Vorhandenseins geeigneter und ungeeigneter Gebiete für das Leben von Beutetieren) sowie die Möglichkeit der Migration von Beutetieren und Raubtieren (unter Beibehaltung). ein gewisser Vorteil der Beute in der Geschwindigkeit dieses Prozesses). Mit anderen Worten, ein Raubtier kann die eine oder andere lokale Ansammlung von Beutetieren vollständig ausrotten, aber einige der Beutetiere haben Zeit, zu wandern und andere lokale Ansammlungen zu bilden. Früher oder später wird das Raubtier auch neue lokale Ansammlungen erreichen, aber in der Zwischenzeit wird die Beute Zeit haben, sich an anderen Orten niederzulassen (auch dort, wo sie zuvor lebte, dann aber ausgerottet wurde).
Ähnliches wie Huffaker im Experiment beobachtete, passiert auch unter natürlichen Bedingungen. Zum Beispiel der Kaktusmotten-Schmetterling (Cactoblastis Kakteen), Nach Australien gebracht, reduzierte sich die Anzahl der Feigenkakteen erheblich, vernichtete sie jedoch nicht vollständig, gerade weil sich der Kaktus etwas schneller ausbreiten konnte. An den Stellen, an denen der Feigenkaktus vollständig ausgerottet ist, kommt die Motte nicht mehr vor. Wenn der Feigenkaktus also nach einiger Zeit wieder hier eindringt, kann er eine gewisse Zeit lang wachsen, ohne dass die Gefahr besteht, dass er von der Motte zerstört wird. Mit der Zeit taucht die Motte jedoch auch hier wieder auf und zerstört durch schnelle Vermehrung den Feigenkaktus.
Wenn man über Raubtier-Beute-Schwankungen spricht, kann man nicht umhin, die zyklischen Veränderungen in der Anzahl der Hasen und Luchse in Kanada zu erwähnen, die auf der Grundlage der Statistiken über die Pelzjagd der Hudson Bay Company vom Ende des 18. Jahrhunderts bis zum Beginn des 20. Jahrhunderts verfolgt werden 20. Jahrhundert. Dieses Beispiel wurde oft als klassische Veranschaulichung der Räuber-Beute-Schwankungen angesehen, obwohl wir in Wirklichkeit nur das Populationswachstum des Räubers (Luchs) im Anschluss an das Wachstum der Beutepopulation (Hase) sehen. Der Rückgang der Hasenzahl nach jedem Aufstieg konnte nicht nur durch den erhöhten Druck von Raubtieren erklärt werden, sondern war offenbar mit anderen Faktoren verbunden, vor allem mit dem Mangel an Nahrung Winterzeit. Zu dieser Schlussfolgerung kam insbesondere M. Gilpin, der zu prüfen versuchte, ob diese Daten durch das klassische Lotka-Volterra-Modell beschrieben werden können. Die Testergebnisse zeigten, dass es keine zufriedenstellende Anpassung an das Modell gab, aber seltsamerweise wurde es besser, wenn Raubtier und Beute getauscht wurden, d. h. interpretierte den Luchs als „Beute“ und den Hasen als „Raubtier“. Eine ähnliche Situation spiegelt sich im humorvollen Titel des Artikels wider („Fressen Hasen Luchse?“), der im Wesentlichen sehr ernst ist und in einer seriösen wissenschaftlichen Zeitschrift veröffentlicht wurde.
Häufig ernähren sich Mitglieder einer Art (Population) von Mitgliedern einer anderen Art.
Das Lotka-Volterra-Modell ist ein Modell der gegenseitigen Existenz zweier Populationen vom Typ „Raubtier-Beute“.
Das Raubtier-Beute-Modell wurde erstmals 1925 von A. Lotka entwickelt, der es zur Beschreibung der Dynamik interagierender biologischer Populationen verwendete. Im Jahr 1926 wurden unabhängig von Lotka ähnliche (und komplexere) Modelle vom italienischen Mathematiker V. Volterra entwickelt, der intensiv auf diesem Gebiet forschte Umweltprobleme legte den Grundstein für die mathematische Theorie biologischer Gemeinschaften oder der sogenannten. mathematische Ökologie.
In mathematischer Form hat das vorgeschlagene Gleichungssystem die Form:
Dabei ist x die Anzahl der Beute, y die Anzahl der Raubtiere, t die Zeit und α, β, γ, δ Koeffizienten, die die Wechselwirkungen zwischen Populationen widerspiegeln.
Formulierung des Problems
Stellen Sie sich einen geschlossenen Raum vor, in dem zwei Populationen existieren – Pflanzenfresser („Beute“) und Raubtiere. Man geht davon aus, dass keine Tiere importiert oder exportiert werden und dass genug Nahrung für Pflanzenfresser vorhanden ist. Dann sieht die Gleichung zur Änderung der Anzahl der Opfer (nur Opfer) wie folgt aus:
wobei $α$ die Geburtenrate der Opfer ist,
$x$ ist die Größe der Beutepopulation,
$\frac(dx)(dt)$ ist die Wachstumsrate der Beutepopulation.
Wenn Raubtiere nicht jagen, können sie aussterben, was bedeutet, dass die Gleichung für die Anzahl der Raubtiere (nur Raubtiere) lautet:
Wobei $γ$ die Raubtierverlustrate ist,
$y$ ist die Größe der Raubtierpopulation,
$\frac(dy)(dt)$ ist die Wachstumsrate der Raubtierpopulation.
Wenn Raubtiere und Beute aufeinander treffen (die Häufigkeit der Begegnungen ist direkt proportional zum Produkt), können Raubtiere ihre Opfer mit einem Koeffizienten vernichten; gut genährte Raubtiere können mit einem Koeffizienten Nachkommen vermehren. Somit hat das Gleichungssystem des Modells die Form:
Die Lösung des Problems
Lassen Sie uns ein mathematisches Modell der Koexistenz zweier biologischer Populationen vom Typ „Raubtier-Beute“ erstellen.
Lassen Sie zwei biologische Populationen in einer isolierten Umgebung zusammenleben. Die Umwelt ist stationär und bietet einer der Arten – den Opfern – unbegrenzte Mengen an allem, was zum Leben notwendig ist. Eine andere Art, ein Raubtier, lebt ebenfalls stationär, ernährt sich jedoch nur von Beute. Als Raubtiere können Katzen, Wölfe, Hechte und Füchse fungieren, als Opfer Hühner, Hasen, Karausche und Mäuse.
Um genauer zu sein, betrachten wir Katzen als Raubtiere und Hühner als Opfer.
Hühner und Katzen leben also in einem isolierten Raum – einem Bauernhof. Die Umwelt bietet Hühnern unbegrenzt Nahrung, und Katzen fressen nur Hühner. Bezeichnen wir mit
$x$ – Anzahl der Hühner,
$у$ – Anzahl der Katzen.
Mit der Zeit ändert sich die Anzahl der Hühner und Katzen, aber wir betrachten $x$ und $y$ als stetige Funktionen der Zeit t. Nennen wir ein Zahlenpaar $x, y)$ den Zustand des Modells.
Lassen Sie uns herausfinden, wie sich der Zustand des Modells $(x, y).$ ändert
Betrachten wir $\frac(dx)(dt)$ – die Änderungsrate der Anzahl der Hühner.
Wenn es keine Katzen gibt, dann nimmt die Anzahl der Hühner zu, und zwar umso schneller, je mehr Hühner es gibt. Wir gehen davon aus, dass die Abhängigkeit linear ist:
$\frac(dx)(dt) a_1 x$,
$a_1$ ist ein Koeffizient, der nur von den Lebensbedingungen der Hühner abhängt natürliche Sterblichkeit und Fruchtbarkeit.
$\frac(dy)(dt)$ – die Änderungsrate der Anzahl der Katzen (wenn es keine Hühner gibt), hängt von der Anzahl der Katzen y ab.
Wenn es keine Hühner gibt, nimmt die Anzahl der Katzen ab (sie haben kein Futter) und sie sterben aus. Wir gehen davon aus, dass die Abhängigkeit linear ist:
$\frac(dy)(dt) - a_2 y$.
In einem Ökosystem wird die Änderungsrate der Anzahl jeder Art ebenfalls als proportional zu ihrer Menge angesehen, jedoch nur mit einem Koeffizienten, der von der Anzahl der Individuen einer anderen Art abhängt. Bei Hühnern nimmt dieser Koeffizient also mit zunehmender Katzenzahl ab und bei Katzen steigt er mit zunehmender Hühnerzahl. Wir gehen außerdem davon aus, dass die Abhängigkeit linear ist. Dann erhalten wir ein System von Differentialgleichungen:
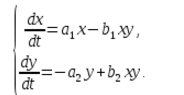
Dieses Gleichungssystem wird Volterra-Lotka-Modell genannt.
a1, a2, b1, b2 – numerische Koeffizienten, die als Modellparameter bezeichnet werden.
Wie Sie sehen, wird die Art der Zustandsänderung des Modells (x, y) durch die Werte der Parameter bestimmt. Durch die Änderung dieser Parameter und die Lösung des Modellgleichungssystems ist es möglich, die Muster der Veränderungen im Zustand des Ökosystems zu untersuchen.
Mit Hilfe MATLAB-Programme Das Lotka-Volterra-Gleichungssystem wird wie folgt gelöst:
In Abb. 1 zeigt die Lösung des Systems. Abhängig von Anfangsbedingungen Die Lösungen sind unterschiedlich, was antworten sie? verschiedene Farben Flugbahnen.
In Abb. 2 präsentiert die gleichen Lösungen, jedoch unter Berücksichtigung der Zeitachse t (d. h. es besteht eine Abhängigkeit von der Zeit).
Zurück in den 20er Jahren. A. Lotka und etwas später, unabhängig von ihm, V. Volter schlugen mathematische Modelle vor, die konjugierte Schwankungen in der Anzahl von Raubtieren und Beutetieren beschreiben.
Das Modell besteht aus zwei Komponenten:
C – Anzahl der Raubtiere; N – Anzahl der Opfer;
Nehmen wir an, dass die Beutepopulation ohne Raubtiere exponentiell wächst: dN/dt = rN. Aber Beute wird von Raubtieren mit einer Geschwindigkeit zerstört, die durch die Häufigkeit der Begegnungen zwischen Raubtier und Beute bestimmt wird, und die Häufigkeit der Begegnungen nimmt zu, wenn die Anzahl der Raubtiere (C) und der Beute (N) zunimmt. Die genaue Anzahl der angetroffenen und erfolgreich gefressenen Beute hängt von der Effizienz ab, mit der das Raubtier die Beute findet und fängt, d. h. von a’ – „Sucheffizienz“ oder „Angriffshäufigkeit“. Somit ist die Häufigkeit „erfolgreicher“ Begegnungen zwischen Raubtier und Beute und damit die Fressrate der Opfer gleich a’СN und im Allgemeinen: dN/dt = rN – a’CN (1*).
Mangels Nahrung verlieren einzelne Raubtiere an Gewicht, verhungern und sterben. Nehmen wir an, dass im betrachteten Modell die Populationsgröße eines Raubtiers bei fehlender Nahrung aufgrund von Hunger exponentiell abnimmt: dC/dt = - qC, wobei q die Sterblichkeit ist. Der Tod wird durch die Geburt neuer Individuen in einem Tempo kompensiert, das in diesem Modell vermutlich von zwei Umständen abhängt:
1) Rate des Lebensmittelverbrauchs, a’CN;
2) die Effizienz (f), mit der diese Nahrung an die Nachkommen des Raubtiers weitergegeben wird.
Somit ist die Fruchtbarkeit des Raubtiers gleich fa’CN und im Allgemeinen gilt: dC/dt = fa’CN – qC (2*). Die Gleichungen 1* und 2* bilden das Lotka-Wolter-Modell. Die Eigenschaften dieses Modells können untersucht, Linienisoklinen konstruiert werden, die einer konstanten Populationsgröße entsprechen, und mit Hilfe solcher Isoklinen kann das Verhalten interagierender Räuber-Beute-Populationen bestimmt werden.
Im Falle einer Beutepopulation: dN/dt = 0, rN = a’CN oder C = r/a’. Weil r und a’ = const, die Isokline für das Opfer ist die Linie, für die der Wert von C konstant ist:
Bei geringer Raubtierdichte (C) nimmt die Beutezahl (N) zu, im Gegenteil ab.
Ähnliches gilt für Raubtiere (Gleichung 2*) mit dC/dt = 0, fa’CN = qC oder N = q/fa’, d. h. Eine Isokline für ein Raubtier ist eine Linie, entlang der N konstant ist: Bei hohen Beutedichten nimmt die Populationsgröße des Raubtiers zu und bei niedrigen Dichten ab.
Ihre Anzahl unterliegt unbegrenzten konjugierten Schwankungen. Wenn die Anzahl der Beutetiere groß ist, nimmt die Anzahl der Raubtiere zu, was dazu führt, dass der Druck der Raubtiere auf die Beutepopulation zunimmt und dadurch deren Anzahl abnimmt. Dieser Rückgang führt wiederum zu einer Einschränkung der Nahrungsaufnahme der Raubtiere und zu einem Rückgang ihrer Zahl, was zu einer Abschwächung des Raubtierdrucks und einer Zunahme der Beutezahl führt, was wiederum zu einer Zunahme der Raubtierpopulation führt , usw.
Populationen führen auf unbestimmte Zeit denselben Schwingungszyklus durch, bis einige Äußerer Einfluss werden ihre Zahlen nicht ändern, woraufhin die Populationen neue Zyklen unbegrenzter Schwankungen durchlaufen. Tatsächlich verändert sich die Umwelt ständig und die Populationen werden ständig neue Niveaus erreichen. Damit die Schwingungszyklen, die eine Population ausführt, regelmäßig sind, müssen sie stabil sein: Wenn ein äußerer Einfluss das Bevölkerungsniveau verändert, müssen sie zum ursprünglichen Zyklus tendieren. Solche Zyklen werden stabile Grenzzyklen genannt.
Das Lotka-Wolter-Modell ermöglicht es uns, den Haupttrend in der Räuber-Beute-Beziehung aufzuzeigen, der sich im Auftreten von Schwankungen in der Beutepopulation ausdrückt, begleitet von Schwankungen in der Räuberpopulation. Der Hauptmechanismus solcher Schwankungen ist die Zeitverzögerung, die der Abfolge von Zuständen von einer hohen Zahl an Beutetieren zu einer großen Zahl an Raubtieren, dann zu einer geringen Zahl an Beutetieren und einer geringen Zahl an Raubtieren und dann zu einer hohen Zahl an Beutetieren innewohnt. usw.
5) BEVÖLKERUNGSSTRATEGIEN VON RÄUBIEREN UND PRIMATEN
Die „Raubtier-Beute“-Beziehung stellt die Verbindungen im Prozess der Übertragung von Materie und Energie von Phytophagen auf Zoophagen oder von Raubtieren niedrigerer Ordnung auf Raubtiere dar Auftrag von oben. Von Aufgrund der Art dieser Beziehungen werden drei Arten von Raubtieren unterschieden:
A) Sammler. Das Raubtier sammelt kleine, ziemlich zahlreiche mobile Opfer. Diese Art der Raubjagd ist typisch für viele Vogelarten (Regenpfeifer, Finken, Pieper usw.), die ihre Energie nur für die Suche nach Opfern aufwenden;
B) wahre Raubtiere. Das Raubtier verfolgt die Beute und tötet sie;
V) Pastoralisten. Diese Raubtiere nutzen wiederholt Beute, zum Beispiel Bremsen oder Bremsen.
Die Strategie zur Nahrungsbeschaffung bei Raubtieren zielt darauf ab, die Energieeffizienz der Ernährung sicherzustellen: Der Energieaufwand für die Nahrungsbeschaffung sollte geringer sein als die Energie, die bei der Nahrungsaufnahme gewonnen wird.
Wahre Raubtiere sind geteilt in
„Schneider“, die sich von reichlich vorhandenen Ressourcen ernähren (einschließlich Planktonfischen und sogar Bartenwal) und „Jäger“, die weniger reichlich Nahrung erhalten. Wiederum
„Jäger“ werden in „Hinterhalte“ unterteilt, die auf Beute lauern (z. B. Hecht, Habicht, Katze, Fangschreckenkäfer), „Sucher“ (insektenfressende Vögel) und „Verfolger“. Für die letztere Gruppe erfordert die Nahrungssuche keinen großen Energieaufwand, wohl aber viel Energie, um die Beute (Löwen in den Savannen) in Besitz zu nehmen. Einige Raubtiere können jedoch Elemente der Strategie verschiedener Jagdoptionen kombinieren.
Wie bei der „Phytophagen-Pflanze“-Beziehung kommt es in der Natur nicht zu einer Situation, in der alle Opfer von Raubtieren gefressen werden, was letztendlich zu ihrem Tod führt. Ökologisches Gleichgewicht Die Verbindung zwischen Raubtieren und Beutetieren wird durch spezielle Mechanismen aufrechterhalten, wodurch das Risiko einer vollständigen Vernichtung der Opfer verringert wird. Opfer können also:
Lauf vor einem Raubtier davon. In diesem Fall nimmt durch Anpassungen die Mobilität sowohl von Opfern als auch von Raubtieren zu, was besonders typisch für Steppentiere ist, die sich vor ihren Verfolgern nirgendwo verstecken können;
Erwerben Sie schützende Farben (stellen Sie sich vor, Blätter oder Zweige zu sein) oder umgekehrt grelle Farbe, N.: rot, warnt Raubtiere vor dem bitteren Geschmack. Es ist bekannt, dass sich die Farbe eines Hasen ändert andere Zeiten Jahr, was ihm ermöglicht, sich im Sommer im Gras und im Winter vor dem Hintergrund des weißen Schnees zu tarnen. Adaptive Veränderung Die Färbung kann in verschiedenen Stadien der Ontogenese auftreten: Robbenjunge sind weiß (Schneefarbe) und ausgewachsene Tiere sind schwarz (Felsküstenfarbe);
Verteilen Sie sich in Gruppen, was das Suchen und Fangen für das Raubtier energieintensiver macht;
Verstecke dich in Notunterkünften;
Gehen Sie zu aktiven Abwehrmaßnahmen über (Pflanzenfresser mit Hörnern, Stachelfische), manchmal auch zu gemeinsamen (Moschusochsen können eine „Rundumverteidigung“ gegen Wölfe usw. übernehmen).
Im Gegenzug entwickeln Raubtiere nicht nur die Fähigkeit, Beute schnell zu verfolgen, sondern auch einen Geruchssinn, der es ihnen ermöglicht, den Standort der Beute anhand des Geruchs zu bestimmen. Viele Raubtierarten reißen die Höhlen ihrer Opfer auf (Füchse, Wölfe).
Gleichzeitig tun sie selbst alles, um ihre Anwesenheit nicht zu entdecken. Dies erklärt die Sauberkeit kleiner Katzen, die viel Zeit mit dem Toilettengang und dem Vergraben von Exkrementen verbringen, um Gerüche zu beseitigen. Raubtiere tragen „Tarngewänder“ (Streifen von Hechten und Barschen, wodurch sie im Dickicht von Makrophyten, Streifen von Tigern usw. weniger auffallen).
Kompletter Schutz von Raubtieren aller Individuen in Populationen von Beutetieren kommt es ebenfalls nicht vor, da dies nicht nur zum Tod hungernder Raubtiere, sondern letztlich zu einer Katastrophe der Beutepopulationen führen würde. Gleichzeitig verschlechtert sich bei Abwesenheit oder Abnahme der Populationsdichte von Raubtieren der Genpool der Beutepopulation (kranke und alte Tiere bleiben erhalten) und durch einen starken Anstieg ihrer Zahl wird die Nahrungsversorgung untergraben.
Aus diesem Grund wird der Effekt der Abhängigkeit der Populationsgrößen von Beutetieren und Raubtieren – ein Pulsieren der Beutepopulationsgröße, gefolgt von einem Pulsieren der Raubtierpopulationsgröße mit einer gewissen Verzögerung („Lotka-Volterra-Effekt“) – selten beobachtet .
Es stellt sich ein recht stabiles Verhältnis zwischen der Biomasse von Raubtieren und Beutetieren ein. So liefert R. Ricklefs Daten, dass das Verhältnis von Raubtier- und Beutebiomasse zwischen 1:150 und 1:300 liegt. In verschiedenen Ökosystemen der gemäßigten Zone der Vereinigten Staaten kommen auf einen Wolf 300 kleine Weißwedelhirsche (Gewicht 60 kg), 100 große Wapitihirsche (Gewicht 300 kg) oder 30 Elche (Gewicht 350). Das gleiche Muster wurde in Savannen gefunden.
Durch die intensive Ausbeutung phytophager Populationen werden Raubtiere häufig aus Ökosystemen ausgeschlossen (in Großbritannien gibt es beispielsweise Rehe und Hirsche, aber keine Wölfe; in künstlichen Stauseen, in denen Karpfen und andere Teichfische gezüchtet werden, gibt es keine Hechte). In diesem Fall übernimmt die Person selbst die Rolle des Raubtiers und entfernt einen Teil der Individuen der Phytophagenpopulation.
Eine besondere Variante der Prädation wird bei Pflanzen und Pilzen beobachtet. Im Pflanzenreich gibt es etwa 500 Arten, die in der Lage sind, Insekten zu fangen und sie mithilfe proteolytischer Enzyme teilweise zu verdauen. Raubpilze bilden Fangvorrichtungen in Form kleiner ovaler oder kugelförmiger Köpfe, die sich auf kurzen Zweigen des Myzels befinden. Die gebräuchlichste Art von Falle sind jedoch selbstklebende dreidimensionale Netze, die aus bestehen große Zahl Ringe, die durch verzweigte Hyphen entstehen. Raubpilze können ziemlich große Tiere, wie zum Beispiel Spulwürmer, fangen. Nachdem sich der Wurm in den Hyphen verfangen hat, wachsen diese in den Körper des Tieres hinein und füllen ihn schnell aus.
1.Konstante und günstige Temperatur- und Luftfeuchtigkeitswerte.
2. Fülle an Nahrungsmitteln.
3. Schutz vor schädlichen Faktoren.
4. Aggressiv chemische Zusammensetzung Lebensraum (Verdauungssäfte).
1. Das Vorhandensein von zwei Lebensräumen: Die Umgebung erster Ordnung ist der Wirtsorganismus, die Umgebung zweiter Ordnung ist die äußere Umgebung.
Computermodell „Raubtier-Opfer“
Kazachkov Igor Alekseevich 1, Guseva Elena Nikolaevna 2
1 Staatliche Technische Universität Magnitogorsk, benannt nach. G.I. Nosova, Institut für Bauwesen, Architektur und Kunst, Studentin im 5. Jahr
2 Staatliche Technische Universität Magnitogorsk, benannt nach. G.I. Nosov, Institut für Energie und automatisierte Systeme, Kandidat der Pädagogischen Wissenschaften, außerordentlicher Professor der Fakultät für Wirtschaftsinformatik und Informationstechnologien
Anmerkung
Dieser Artikel ist einem Überblick über das Computermodell „Raubtier-Beute“ gewidmet. Die durchgeführte Forschung legt nahe, dass Umweltmodellierung eine große Rolle in der Umweltforschung spielt. Dieses Problem ist vielfältig.
Computermodell „Raubtier-Opfer“
Kazatchkov Igor Alekseevich 1, Guseva Elena Nikolaevna 2
1 Staatliche Technische Universität Nosov Magnitogorsk, Institut für Bauingenieurwesen, Architektur und Kunst, Student des 5. Studiengangs
2 Staatliche Technische Universität Nosov Magnitogorsk, Institut für Energietechnik und automatisierte Systeme, Doktor der Pädagogik, außerordentlicher Professor der Abteilung für Wirtschaftsinformatik und Informationstechnologien
Abstrakt
Dieser Artikel gibt einen Überblick über das Computermodell „Raubtier-Opfer“. Die Studie legt nahe, dass Umweltsimulationen eine große Rolle bei der Untersuchung der Umwelt spielen. Dieses Problem ist vielfältig.
Ökologische Modellierung wird zur Untersuchung unserer Umwelt eingesetzt. Mathematische Modelle werden in Fällen verwendet, in denen keine natürliche Umgebung und keine natürlichen Objekte vorhanden sind. Sie helfen dabei, den Einfluss verschiedener Faktoren auf das untersuchte Objekt vorherzusagen. Diese Methodeübernimmt die Aufgabe, die gewonnenen Ergebnisse zu überprüfen, zu konstruieren und zu interpretieren. Basierend auf solchen Formen befasst sich die Umweltmodellierung mit der Bewertung von Veränderungen in der Umwelt um uns herum.
IN momentanÄhnliche Formen werden verwendet, um die Umwelt um uns herum zu untersuchen, und wenn es notwendig ist, einen ihrer Bereiche zu untersuchen, verwenden sie sie Mathe-Modellierung. Dieses Modell ermöglicht es, den Einfluss bestimmter Faktoren auf den Untersuchungsgegenstand vorherzusagen. Der „Raubtier-Beute“-Typ wurde einst von Wissenschaftlern wie T. Malthus (Malthus 1798, Malthus 1905), Verhulst (Verhulst 1838), Pearl (Pearl 1927, 1930) sowie A. Lotka ( Lotka 1925, 1927) und V. Volterra (Volterra 1926) reproduzieren das periodische Schwingungsregime, das als Ergebnis interspezifischer Wechselwirkungen in der Natur entsteht.
Eine der wichtigsten Erkenntnismethoden ist die Modellierung. Zusätzlich zu der Tatsache, dass es Veränderungen vorhersagen kann, die in auftreten Umfeld, hilft auch dabei, den optimalen Weg zur Lösung des Problems zu finden. Mathematische Modelle werden in der Ökologie seit langem verwendet, um Muster und Trends in der Entwicklung von Populationen zu ermitteln und dabei zu helfen, das Wesentliche von Beobachtungen hervorzuheben. Das Layout kann als Muster dienen Verhalten, Objekt.
Bei der Nachbildung von Objekten in der mathematischen Biologie werden Vorhersagen verschiedener Systeme verwendet, besondere Individualitäten von Biosystemen werden berücksichtigt: die innere Struktur des Individuums, Lebenserhaltungsbedingungen, Konstanz Ökosysteme, wodurch die lebenswichtige Aktivität der Systeme erhalten bleibt.
Das Aufkommen der Computermodellierung hat die Grenzen der Forschungsmöglichkeiten erheblich erweitert. Die Möglichkeit der multilateralen Umsetzung schwieriger Formen, die keine analytische Untersuchung zulassen, ist entstanden, ebenso wie die Simulationsmodellierung.
Betrachten wir, was ein Modellierungsobjekt ist. „Das Objekt ist ein geschlossener Lebensraum, in dem eine Interaktion zwischen zwei biologischen Populationen stattfindet: Raubtieren und Beutetieren. Der Prozess des Wachstums, des Aussterbens und der Fortpflanzung findet statt direkt auf der Oberfläche des Lebensraums. Die Beute ernährt sich von den Ressourcen, die in der Umwelt vorhanden sind, während die Raubtiere sich von der Beute ernähren. In diesem Fall können Nahrungsressourcen entweder erneuerbar oder nicht erneuerbar sein.
Im Jahr 1931 leitete Vito Volterra die folgenden Gesetze der Räuber-Beute-Beziehung ab.
Das Gesetz des periodischen Zyklus – der Prozess der Beutevernichtung durch ein Raubtier führt oft zu periodischen Schwankungen der Populationsgröße beider Arten, die nur von der Wachstumsrate der Fleischfresser und Pflanzenfresser und vom anfänglichen Verhältnis ihrer Zahlen abhängen.
Gesetz der Durchschnittserhaltung – Durchschnittszahl jedes Typs ist unabhängig davon konstant Einstiegslevel, vorausgesetzt, dass die spezifischen Raten des Bevölkerungswachstums sowie die Effizienz der Raubtiere konstant sind.
Das Gesetz der Verletzung von Durchschnittswerten – wenn beide Arten proportional zu ihrer Zahl reduziert werden, nimmt die durchschnittliche Populationsgröße der Beute zu und die der Raubtiere ab.
Beim Räuber-Beute-Modell handelt es sich um eine besondere Beziehung zwischen einem Räuber und seiner Beute, von der beide profitieren. Es überleben die gesündesten und am besten an die Umweltbedingungen angepassten Individuen, d. h. All dies geschieht dank natürliche Auslese. In einer Umgebung, in der es keine Möglichkeit zur Fortpflanzung gibt, Das Raubtier wird früher oder später die Population der Beute vernichten, wodurch diese selbst aussterben wird.“
Auf der Erde gibt es viele Lebewesen, die unter günstigen Bedingungen die Zahl ihrer Verwandten enorm vermehren. Diese Fähigkeit nennt man: das biotische Potenzial einer Art, d.h. eine Zunahme der Zahl einer Art über einen bestimmten Zeitraum. Jede Art hat beispielsweise ihr eigenes biotisches Potenzial große Arten Organismen pro Jahr können nur um das 1,1-fache zunehmen, wiederum Organismen kleinerer Arten wie Krebstiere usw. können ihr Aussehen um das 1030-fache steigern, Bakterien jedoch immer noch mehr. In jedem dieser Fälle wird die Bevölkerung exponentiell wachsen.
Exponentielles Bevölkerungswachstum ist ein geometrischer Verlauf des Bevölkerungswachstums. Diese Fähigkeit kann im Labor bei Bakterien und Hefen beobachtet werden. Unter Nicht-Laborbedingungen lässt sich ein exponentielles Wachstum am Beispiel von Heuschrecken oder anderen Insektenarten beobachten. Eine solche Zunahme der Artenzahl ist dort zu beobachten, wo es praktisch keine Feinde gibt und es mehr als genug Nahrung gibt. Schließlich kam es zu einer Zunahme der Arten, nachdem die Zahl für kurze Zeit zugenommen hatte, und das Populationswachstum begann zu sinken.
Betrachten wir ein Computermodell der Säugetierreproduktion am Beispiel des Lotka-Volterra-Modells. Lassen In einem bestimmten Gebiet gibt es zwei Arten von Tieren: Hirsche und Wölfe. Mathematisches Modell der Bevölkerungsveränderungen im Modell Tabletts-Volterra:
Die anfängliche Anzahl der Opfer beträgt xn, die Anzahl der Raubtiere beträgt yn.
Modellparameter:
P1 – Wahrscheinlichkeit, einem Raubtier zu begegnen,
P2 – Wachstumskoeffizient der Raubtiere auf Kosten der Beute,
d – Sterblichkeitsrate der Raubtiere,
a ist der Koeffizient der Zunahme der Opferzahl.
In der Trainingsaufgabe wurden folgende Werte festgelegt: Die Anzahl der Hirsche betrug 500, die Anzahl der Wölfe betrug 10, die Wachstumsrate der Hirsche betrug 0,02, die Wachstumsrate der Wölfe betrug 0,1, die Wahrscheinlichkeit, einem Raubtier zu begegnen, betrug 0,0026, die Wachstumsrate der Raubtiere auf Kosten der Beute betrug 0,000056. Die Daten werden für 203 Jahre berechnet.
Wir erforschen den Einfluss der Koeffizient der Opferzunahme für die Entwicklung zweier Populationen, die übrigen Parameter bleiben unverändert. In Schema 1 wird eine Zunahme der Beutezahl und dann, mit einer gewissen Verzögerung, eine Zunahme der Raubtiere beobachtet. Dann schlagen die Raubtiere die Opfer nieder, die Zahl der Opfer sinkt stark und in der Folge nimmt die Zahl der Raubtiere ab (Abb. 1).

Abbildung 1. Bevölkerungsgröße mit niedrigen Geburtenraten unter den Opfern
Lassen Sie uns die Änderung im Modell analysieren, indem wir die Geburtenrate des Opfers um a = 0,06 erhöhen. In Diagramm 2 sehen wir einen zyklischen Oszillationsprozess, der im Laufe der Zeit zu einem Anstieg der Zahlen beider Populationen führt (Abb. 2).

Abbildung 2. Bevölkerungsgröße bei durchschnittlicher Geburtenrate der Opfer
Betrachten wir, wie sich die Bevölkerungsdynamik bei einem hohen Wert der Geburtenrate des Opfers a=1,13 ändert. In Abb. 3 Es kommt zu einem starken Anstieg der Bestände beider Populationen, gefolgt vom Aussterben von Beutetieren und Raubtieren. Dies ist auf die Tatsache zurückzuführen, dass die Beutepopulation so stark zugenommen hat, dass die Ressourcen zur Neige gegangen sind und die Beutetiere ausgestorben sind. Das Aussterben der Raubtiere ist auf die Tatsache zurückzuführen, dass die Zahl der Beutetiere zurückgegangen ist und den Raubtieren die Ressourcen zum Überleben ausgehen.

Abbildung 3. Bevölkerungsgröße mit hohen Geburtenraten unter den Opfern
Basierend auf der Analyse von Computerexperimentdaten können wir den Schluss ziehen, dass Computermodelle es uns ermöglichen, Populationsgrößen vorherzusagen und den Einfluss verschiedener Faktoren auf die Populationsdynamik zu untersuchen. Im obigen Beispiel haben wir das Raubtier-Beute-Modell untersucht, den Einfluss der Geburtenrate von Beutetieren auf die Anzahl von Hirschen und Wölfen. Eine kleine Zunahme der Beutepopulation führt zu einer kleinen Zunahme der Beute, die nach einer gewissen Zeit von Raubtieren vernichtet wird. Eine moderate Zunahme der Beutepopulation führt zu einer Vergrößerung beider Populationen. Ein starker Anstieg der Beutepopulation führt zunächst zu einem schnellen Anstieg der Beutepopulation, dies wirkt sich auf die Zunahme des Raubtierwachstums aus, doch dann zerstören die sich vermehrenden Raubtiere schnell die Hirschpopulation. Dadurch sterben beide Arten aus.



